
Schopenhauer-Gedenkschrift 2016

Gleiches erkennt Gleiches – Annäherungen an alternative Auffassungen zum Farbsehen.
Eckhard Bendin
Ausgehend von Schopenhauers Theorie der partitiv geteilten Tätigkeit der Retina werden Zusammenhänge zwischen den Schwingungsgestalten der eintretenden Lichtstrahlung, den anatomisch-physiologischen Dispositionen und der axialen, paarigen Struktur unserer gegenfarbigen Empfindungen betrachtet. Dabei drängt sich auf, dass neben der Frequenz und der Amplitude der Schwingungen die Bedeutung der den Gestalten inhärenten ‚Schwingungsebenen‘ bisher vielleicht zu Unrecht als dritte, möglicherweise ebenso relevante Größe der Farbdifferenzierung vernachlässigt wurde. Dies wird an Fragestellungen demonstriert, in die auch neuere Überlegungen zur Funktion anatomischer Gegebenheiten der Retina einbezogen werden (Krumeich/ Knülle-Wenzel 1992/ Lauinger 1996 und Franze et al. 1997). Unter der Annahme, dass unsere Farbempfindungen physiologisch geforderte, gegensätzlichpaarige Erscheinungen sind, die eine polar-axial-generierte Struktur bilden, lässt sich der Aufbau der inversen Retina durchaus auch interpretieren als für die Aufnahme komplexer Schwingungsmuster geradezu prädestiniert. Die Interpretation der Müllerzellen als Lichtleiter und an die Rezeptoren gekoppeltes zelluläre Raumgitter lässt bei abgeschwächter Totalreflexion evaneszente Felder zu. Die biaxial und bipolar organisierte Netzhautstruktur, ebenso die hexagonal verdichtete Packung aller Zellschichten lässt auf seine Funktion auch zur Aufnahme komplexer Reiz-Muster in Form von ‚Schwingungsgestalten‘ schließen. Dies legt strukturelle Analogien zwischen den Reizkonfigurationen als ‚Erkanntem‘ und den Verarbeitungsstrukturen als dem ‚Erkennenden‘ nahe und folgt der Erkenntnislehre des Empedokles (492-432 v. Chr.) „Gleiches wird durch Gleiches erkannt.“

Die Basis: Farbe als physiologisch geforderte gegensätzlich-paarige Erscheinung
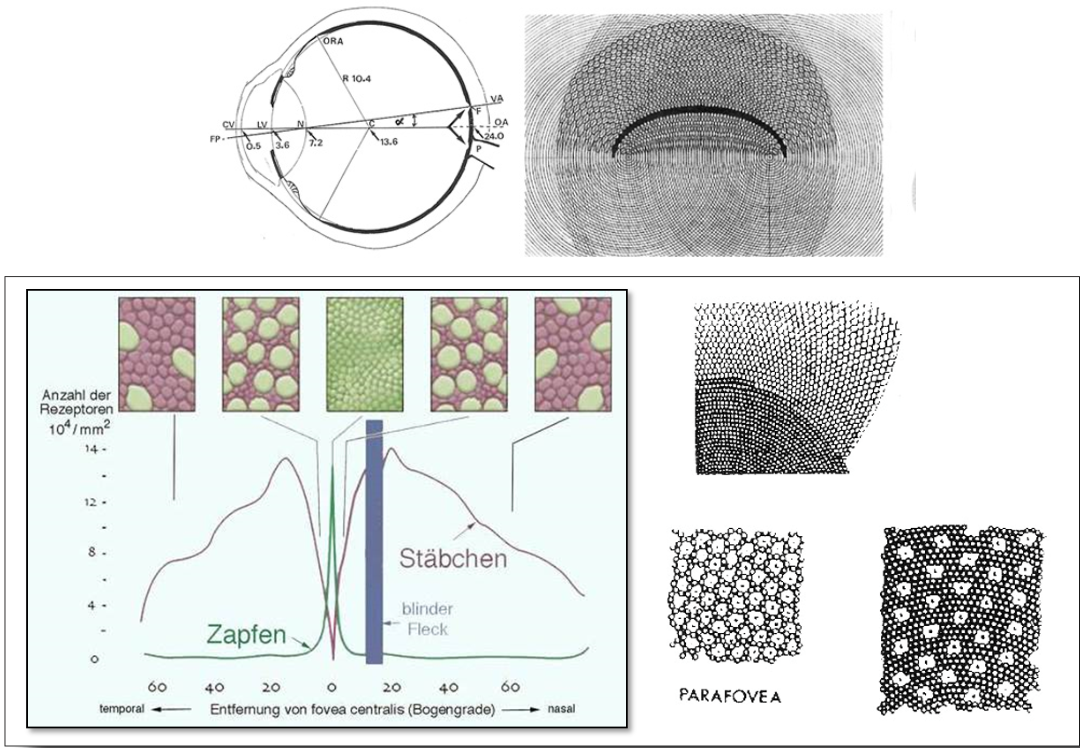
Auf Goethes Erkenntnissen fußend und über sie teilweise hinausgehend interpretierte Schopenhauer 1816 die Gegenfarbenpaare als duale Einheiten einer partitiv geteilten Tätigkeit der Retina (Abb 1). Nach den Untersuchungen Chevreuls 1839 zum Simultankontrast sowie Herings Opponententheorie 1875 wurden antagonistische Prozesse bei der Reizverarbeitung als relevant angenommen. Auf der Grundlage der Dreifarbentheorie von Young/Helmholtz 1855 interpretierte man die Gegenfarben-Prozesse nach der Zonentheorie von v. Kries 1923 (weiterentwickelt durch Hurvich u. Jameson 1957) als relevant für die den Farbrezeptoren nachgeschaltete neuronale Reizverarbeitung in den Ganglienzellen der Netzhaut in Gestalt antagonistischen Erregungs- u- Hemmungsprozesse. (Abb. 2)
Auch ChristineLadd-Franklin stellte 1898 eine evolutionäre Theorie zur Diskussion, die eine stufenartige Rezeptorentwicklung vom Achromatischen zum Chromatischen annahm, 1926 aufgegriffen auch von Schrödinger. Eine ähnliche polar-komplementäre Stufenordnung brachte 1961 der Weizsäcker-Schüler Eckart Heimendahl mit energetischer Begründung vor. Besonders aber soll hier erwähnt werden, dass Wilhelm Wundt in Abgrenzung zu Helmholtz und Hering 1874 eine Stufentheorie entwarf, nach der sich achromatische und chromatische Erregungen in Abhängigkeit von der Amplitude der Lichtschwingungen stufenartig entwickeln und zu der er schlussfolgerte, dass es „überhaupt keine fest begrenzte Zahl farbiger Sehstoffe (gibt), sondern diese bilden eine aus unbestimmt vielen Abstufungen bestehende Reihe.“ [Wundt 1910] (Abb. 3)
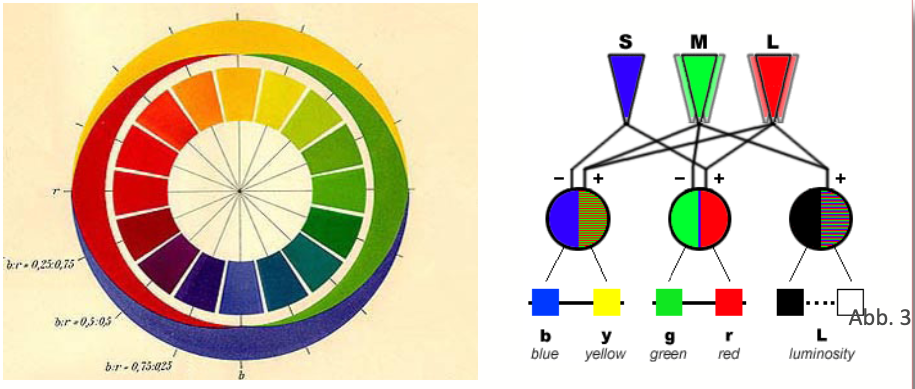

Aus der von Schopenhauer postulierten physiologischen Äquivalenz der Gegenfarben ließ sich eine spezifische Axialität ableiten, wie sie – zunächst 1868 in einer Farbenlehre von Guido Schreiber aufgegriffen und von Adolf Hölzel um 1914 in seinem zwölfteiligen chromatischen Farbkreis bezugnehmend auch auf Wilhelm v. Bezold angeboten- schließlich 1930 von der Hölzel-Schülerin Carry van Biema quantitativ analog zu Schopenhauers Bipartion schematisch vorgestellt wurde. (Abb. 4)

Fragestellung 1: Farberscheinungen als Ausdruck signifikanter Prägung durch inhärente Axialität der Schwingungsgestalten?
In den seltensten Fällen ist unser Auge monochromatischem Licht ausgesetzt. Unser
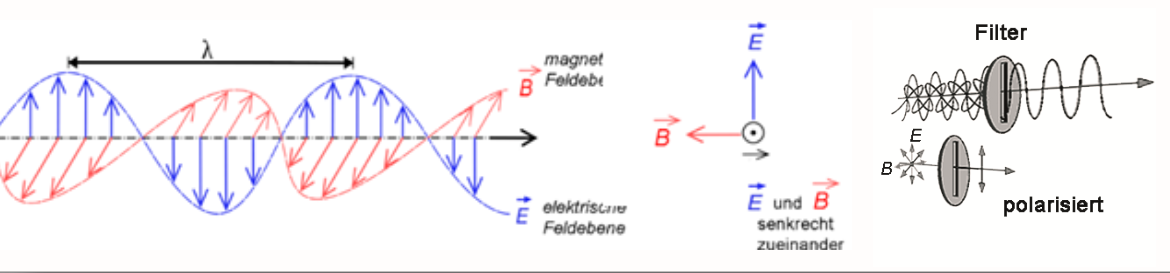
Auge nimmt zumeist zusammengesetzte Lichtstrahlung unterschiedlicher ‚spektraler Verteilung‘ auf, d.h. es handelt sich bei den ‚Resultanten‘ um ‚komplexe Muster‘ aus punktspiegeldrehsymmetrischen Operationen. Elektromagnetische Wellen sind komplexe Schwingungsgestalten (Abb.5). Deren Frequenz und Amplitude können zwar als signifikante Merkmale angesehen werden, nicht zwingend aber als allein hinreichende Ursachen für die Prägung der Farberscheinung [Bendin 1991], denn die ‚Gestalt‘ der Welle wird auch durch die Axialität u. Lage ihrer Schwingungsebenen bestimmt. Diese sind polar-axial ausgerichtet, wobei die elektrischen und magnetischen Feldebenen senkrecht zueinander stehen. Bei Rotationsdispersion an Quarz gibt es z.B. eine auffallend symmetrische Spreizung der Polarisationsfarben nach Bendin 1991/2010 (Abb.6). Auch Stehende Wellen und Interferenzfarben resultieren aus differenzierten Schwingungsgestalten infolge Superposition und Phasenverschiebung (Abb 7).
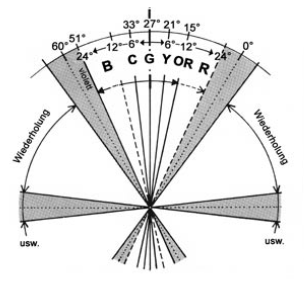


Zudem gibt es eine auffällige Axialität bei Farbfehlsichtigkeiten, welche die Natur der Farberscheinungen als generative Struktur aus polar-axialen Einheiten nahelegt. Die Ermittlung der Fehlsichtigkeiten auf der Grundlage des Farnsworth-Munsell 100 Hue Test ergibt sowohl für die Blau-Gelb-Schwäche (Tritanopie) als auch für die Rot- und Grünschwäche (Protanopie bzw. Deuteranopie) eindeutige axiale Ausfälle.
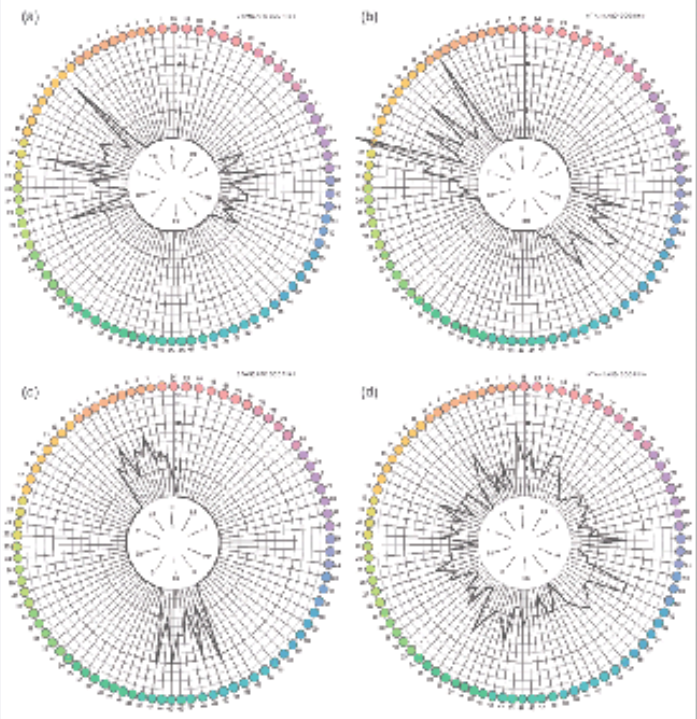
Es gibt also offensichtlich eine Reihe von Entsprechungen, die zur Fragestellung herausfordern: Könnte die der elektromagnetischen Struktur von Schwingungsgestalten inhärente symmetrische Axialität (z.B. Schwingungsebenen bei Polarisationsfarben, Phasenlagen bei Interferenzfarben) nicht auch Einfluß auf die physiologische Signalverarbeitung haben? Dies legt auch die Frage nahe, die Richard Funk in seinem Beitrag stellte: „Warum sollte die Natur die quantenoptischen Phänomene gerade in der Beziehung zu der ausgefeilten Nanostrukturierung der Fotorezeptoren und anderer Zellen nicht ausnutzen?“
Fragestellung 2: Ist eine Rezeption stehender Wellen (z.B. SWA- Colorgramme nach Krumeich/Knülle) als signifikante Schwingungsgestalten vorstellbar?
Es gibt eine auffällige morphologische Analogie zwischen der Nanostruktur der sogen. ‚Schillerschuppen‘ beim ‚Morpho Rhentenor‘ und den ‚Disci‘ (den Außengliedern der Rezeptoren), die eine Rezepzion stehender Wellen in Verbindung mit der inversen Funktion der Retina unter Annahme der Bruchchen Membran als Reflektor durchaus plausibel erscheinen läßt (Abb. 9). Der Abstand der Scheiben der Disci beträgt etwa 55 nm, wodurch sich nach Krumeich/Knülle stehende Wellen bilden und als ‚SWA-Colorgramme‘ registriert werden könnten.
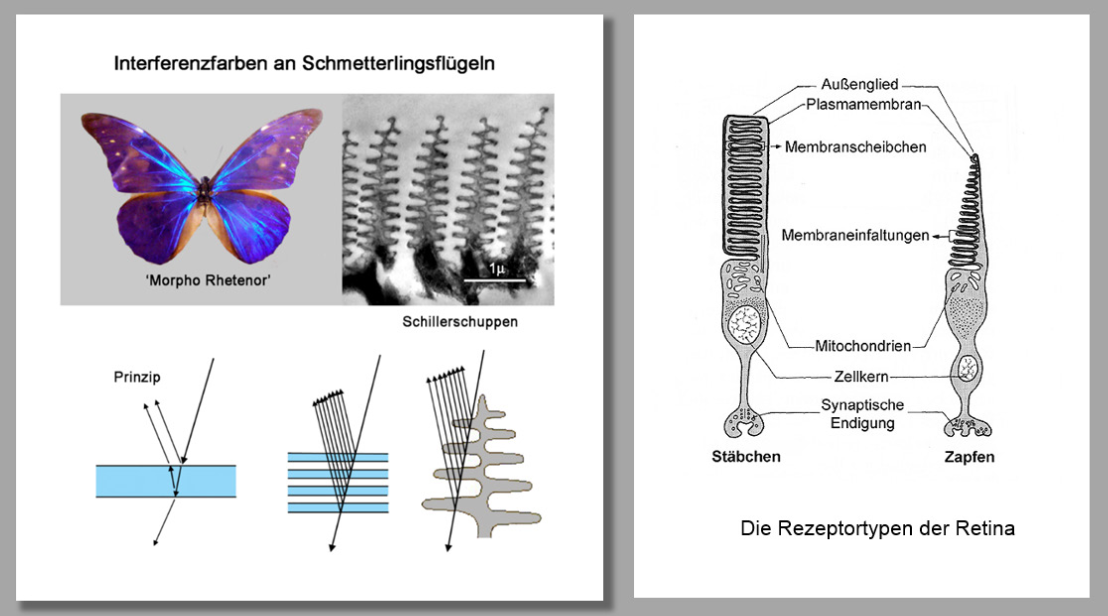
Der inverse Aufbau der Retina erscheint als ideale anatomische Disposition zur Aufnahme komplexer Schwingungsmuster. Nach der Theorie von Krumeich und Knülle ist vorstellbar, dass das durch die verschiedenen Schichten der inversen Retina geführte Licht an der Bruch‘schen Membran reflektiert und von den Disci der Rezeptoren in Gestalt stehender Wellen registriert werden kann. (Abb 10 u. 11)
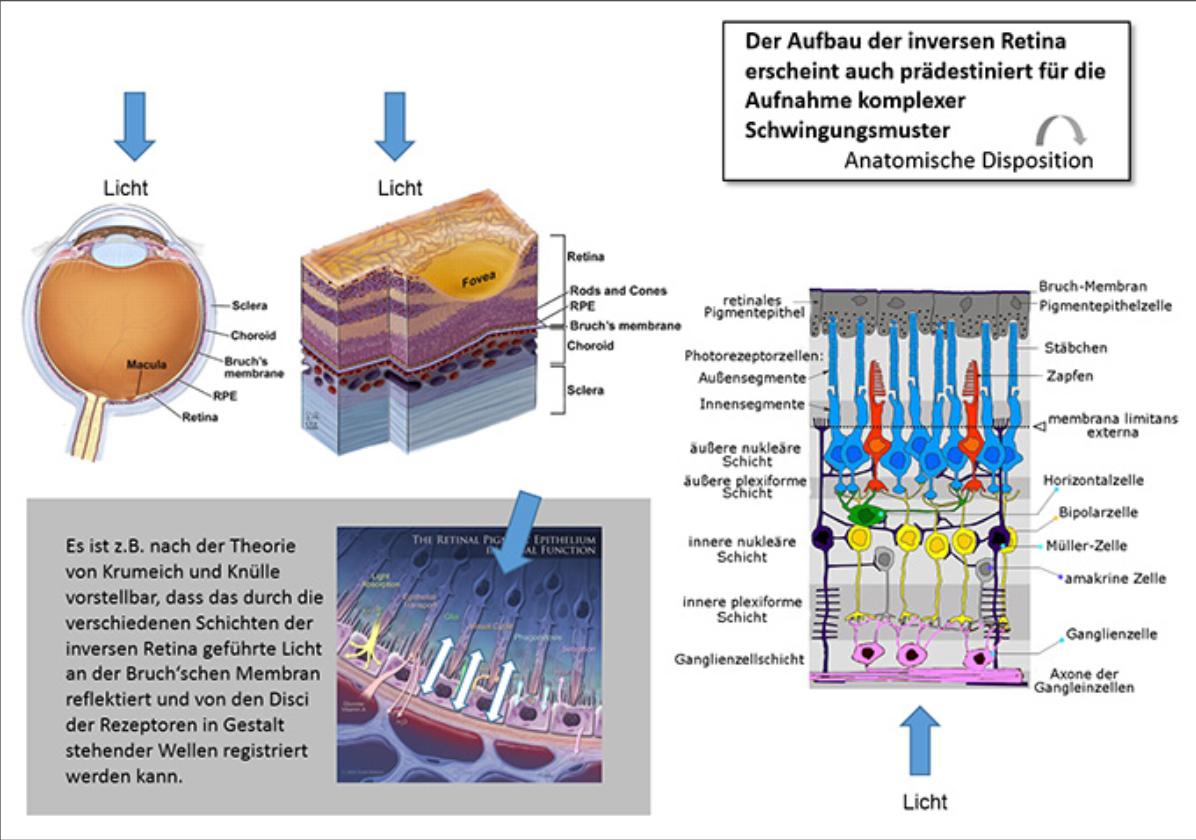
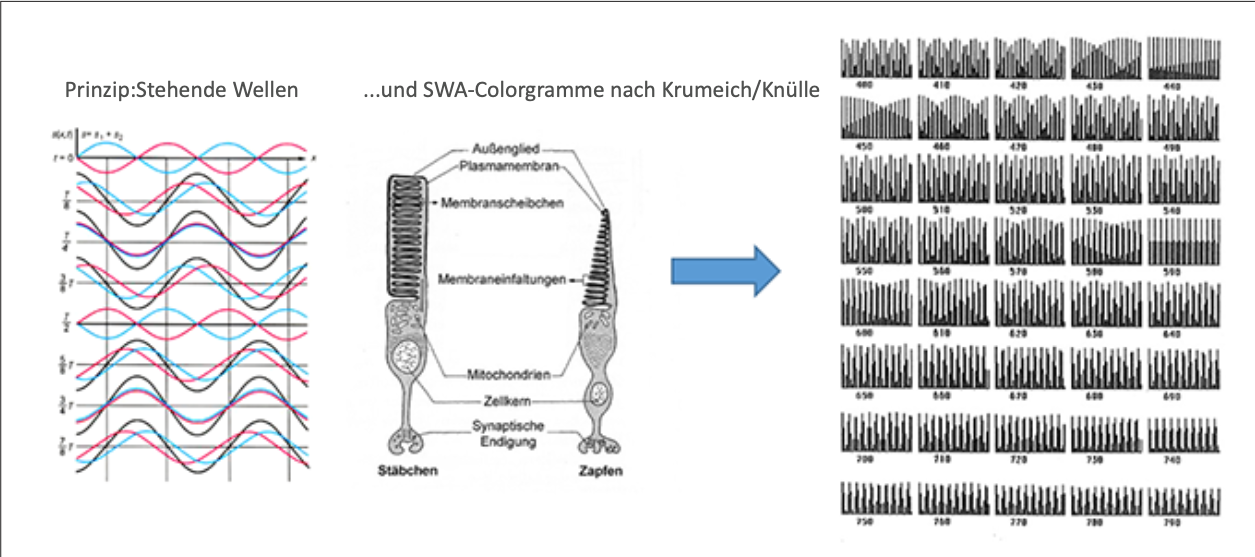
Abb.11: Bestärkt werden kann diese Annahme durch den Nachweis der Funktion der Ganglienzel- len (Müllerzellen) als Lichtleiter. Durch Totalreflexion erfolgt der Lichttransport praktisch ohne Kapazitätsverlust (Abb. 12)
Fragestellung 3: Inwiefern bedeutet die nachgewiesene Funktion der Müllerzellen als Lichtleiter eine Erweiterung der anatomische Disposition?
2007 wurde an Meerschweinchen die Funktion der Müllerzellen als Lichtleiter, als eine
Art „lebendes Glasfaserkabel“, entdeckt [Franze et al. 2007]. Meerschweinchen haben die gleichen Müllerzellen wie der Mensch. Die Müllerzelle durchwächst ein zelluläres ‚Raumgitter‘ [Lauinger 1994/96]. Als eng an die Rezeptoren gekoppeltes Gitter ermöglichen sie eine exakte Datenwiedergabe.
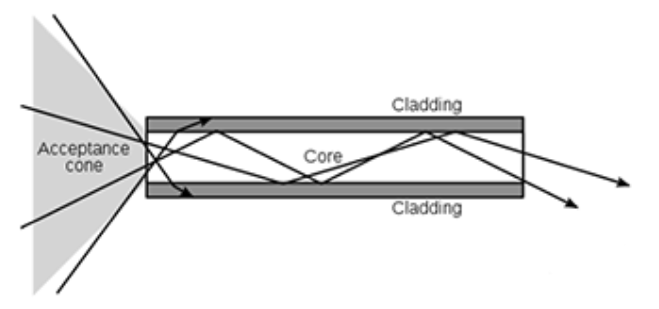

Abb.13:
Müllerzelle und kommerzielle Lichtleiter
a) Das Bild eines winzigen Buchstabens, der zum einen auf einen kommerziellen Lichtleiter, zum anderen auf die Retina eines Meerschweinchens projiziert wird.
b) Wiedergabe des Buchstabens mit kommerziellem Lichtleiter.
c) Wiedergabe des Buchstabens mit den Müllerzellen des Meerschweinchens. Zum Vergleich: Der kleine weiße Strich ist 2 Hundertstelmillimeter lang.
d) Schematische Darstellung des Lichtleiters (Querschnitt)
e) Schematische Darstellung der Müllerzellen in der Retina des Meerschweinchens. Die schwarzen Zwischenräume sind gefüllt mit Nervenzellen.
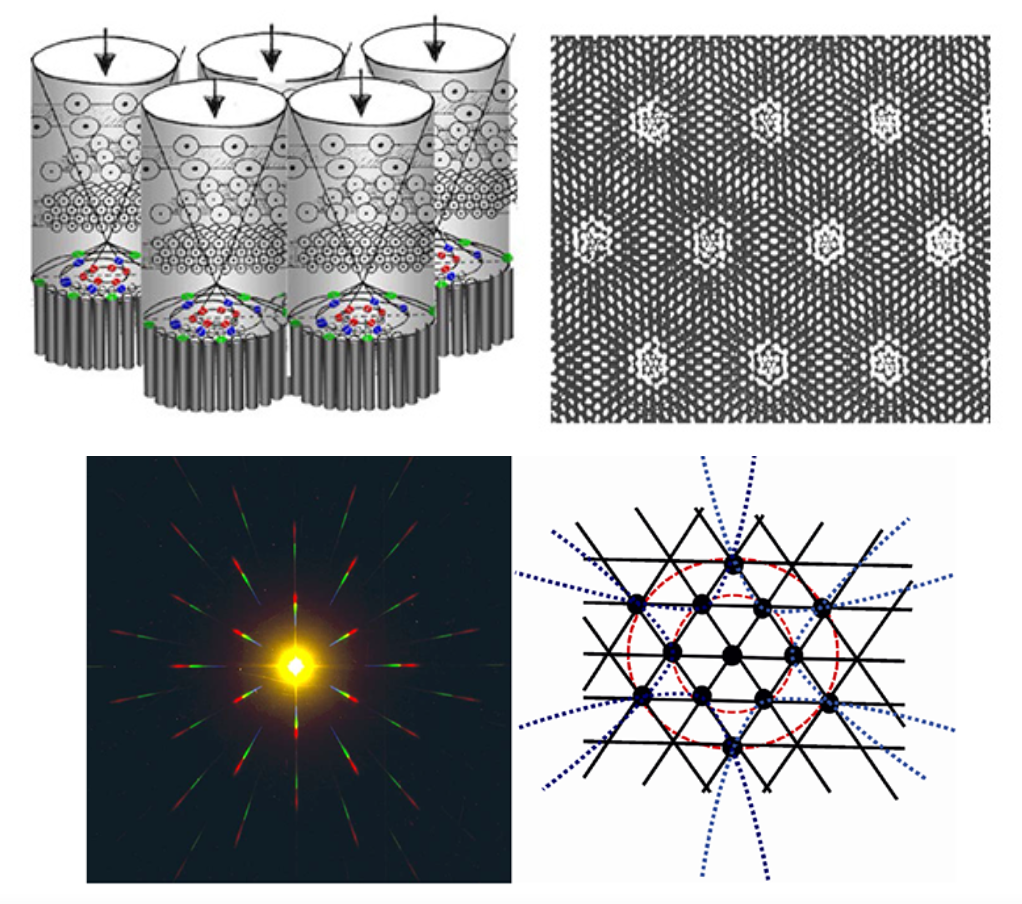
Nach der Auffassung von Lauinger (1994/96) wirkt das zelluläre Raumgitter als Resonator und optischer Korrelator. Die drei zellulären Schichten (Zellkern und Zellplasma als Elemente des Phasengitters) liegen lichtwärts unmittelbar vor der Photorezeptoren (REC), die also im Nahfeld hinter dem zellulären Raumgitter stehen (Abb 14). Die hierarchische Geometrie des retinalen Raumgitters ist hexagonal, mit nach vorn größeren und nach hinten kleineren Gitterkonstanten. Zur Peripherie der Netzhaut hin werden die Gitterkonstanten im vorderen Gitter (ONL) zunehmend größer. Die hexagonale Struktur stellt somit ein polares Gitter dar mit radialkonzentrischer Ordnung auf der Mantelfläche eines Rotationsellipsoids (Innenauge). Lauinger geht von einer “‚optischen Stimmung‘ von lichtbeugenden Gittern und ihrer spektralen Resonanz“ aus. Das bedeutet, dass spezifische Beugungsbilder an hexagonalen Gittern auch die Auslöser der Farbinformationen sein können. „An die Stelle der Summierung von Intensitäten tritt in der Interferenzoptik die Aufteilung bzw. Umverteilung des Lichts in die Polarität von Hell und Dunkel, Licht und Finsternis, Maxima und Minima der Intensitäten. Zwischen beiden Extremen liegt die Gleichgewichtslage von Hell und Dunkel im Mittelwert des kontrastlosen Grau.“[Lauinger 2014]
Fragestellung 4: Sind analog zur optischen Sensorik auch in der Retina evanescente Wechselwirkungen vorstellbar bei abgeschwächter Totalreflexion?
Es stellt sich die Frage, welche Interpretationen das Durchwachsen der Müllerzellen durch das zelluläre Retina-Raumgitter als polare, hexagonale Struktur weiterhin zuläßt? Sind hier z.B. evaneszente Wechselwirkungen denkbar, wie sie in der Sensorik genutzt werden, da auf diese Weise sehr empfindliche Änderungen im evaneszenten Feld gemessen werden
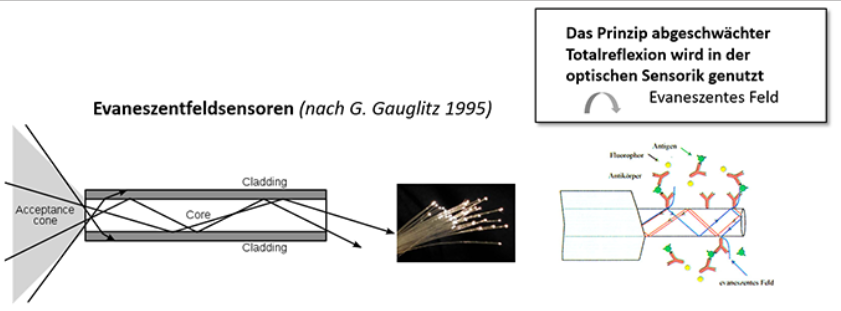
können [nach Gauglitz 1995]? Wenn also Strahlung aus dem bzw. in den Wellenleiter (Müllerzelle) aus- bzw. einkoppeln kann,- abhängig vom Brechungsindex des Mediums, von dem das Gitter umspült wird – ist durchaus vorstellbar, dass nach diesem Prinzip auch in der Retina feinste Differenzierungen registrierbar werden, denkbar auch als interferenzoptische Informationen in Gestalt von Mustern (Abb.16 rechts).

Schemata nach wikipedia.org/wiki/Evaneszenz und nach P. Leuchtmann, ETH Zürich
Fragestellung 5: Wie signifikant sind die axialsymmetrischen Verhältnisse der elektromagnetischen Schwingungen für die Reizverarbeitung?
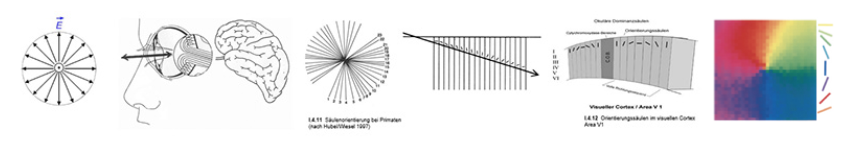
Seit der Beschreibung des differenziellen Interferenzkontrastes sowie der Entwicklung
des Phasenkontrastes in den 50er Jahren durch Normanski bzw. Zernike sind Analogien zwischen Schwingungsgestalt, Phasenmustern u. Beugungsbildern an hexagonalen Gittern stärker von Interesse. Auch zum axialen Schema der ‚Dominanzsäulen‘ im visuellen Cortex/ Area V1 [Hubel/Wiesel 1962/63] gibt es Entsprechungen. Es wäre von Interesse, ob die axialsymmetrische Ausrichtung der Phasenmuster und Beugungsbilder für einen Zusammenhang von Reiz- u. Verarbeitungsstrukturen spricht, der in neuronaler und kortkaler Verarbeitung zur Mannigfaltigkeit der Helligkeits- u. Farbinformationen beiträgt.

In Bezug auf die Reizverarbeitung im visuellen Cortex gibt es auch eine interessante Analogie der in den okulären Dominanzsäulen der Area V1 an Primaten nachgewiesenen richtungsabhängigen Antworten von Neuronen der Sehrinde (von Hubel und Wiesel 1997 – im Kreis dargestellt als eine symmetrisch gespreizte Axialität jener ‚Orientierungssäulen‘ – mit adäquaten Strukturen der Farbtonachsen in Farbtonkreisen mehrerer Autoren (Abb.18).
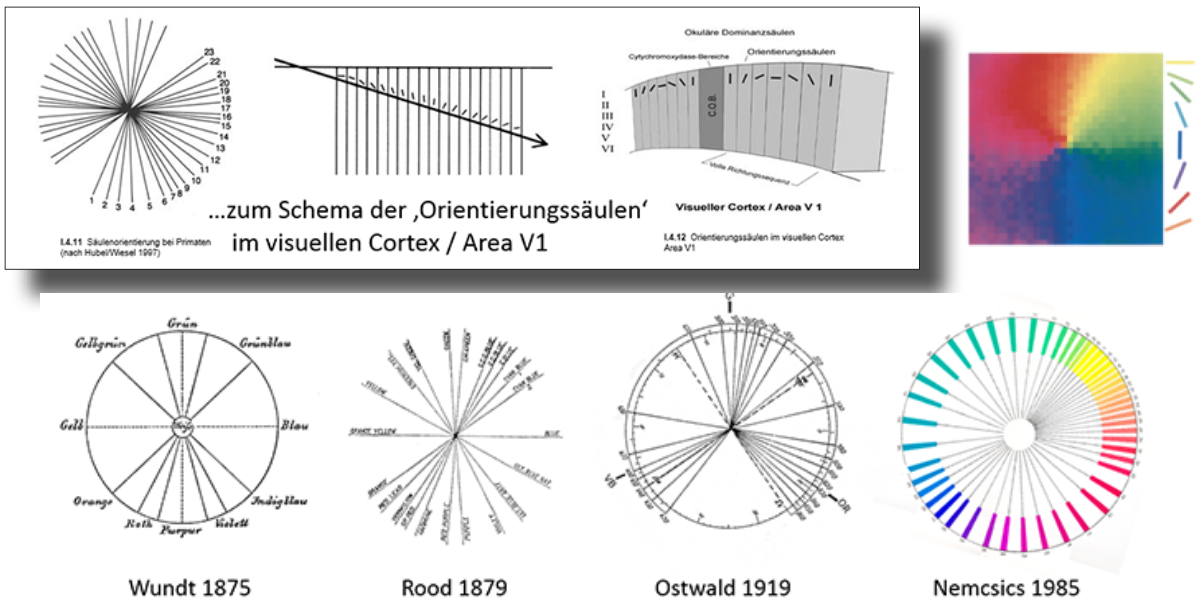
Abb. 18: oben li: Schema der Orientierungssäulen in Area V1 / re: Farb-Mappe zur Orientierungspräferenz nach Purves et.al. 2004 / darunte: Farbkreise von Wundt, Rood, Ostwald u. Nemcsics
Die Annahme wird noch gestützt durch die unmittelbare Kopplung der Orientierungssäulen mit den Cytochromoxydase-Bereichen (COB), denen die Verarbeitung von Farbinformationen zugeschrieben wird. Hinsichtlich der Orientierungspräferenz im visuellen Kortex faßt Lauinger z.B. das Geschehen im Zellgitter der Netzhaut als „spiralartige Windung
des Lichts“ auf und setzt es mit einer entsprechenden Verarbeitung im visuellen Cortex
in Beziehung: „Das interferenzoptische Geschehen gleicht weitgehend einer spiralartigen Windung des Lichts durch das Mehrschichtengitter, wie sie auch der Struktur der ‚orienta- tion pinwheel centers‘ in ocular dominance columns im visual cortex V1 zugrunde zu liegen scheint.“ [Lauinger 2014, S.272]
Fragestellung 6: Unterstützt auch die Rezeptorverteilung in der biaxial und bipolar organisierten Netzhautstruktur die Annahme einer anatomischen Disposition zur Aufnahme komplexer ‚Schwingungsgestalten‘ (Mustern)?
Jedes einzelne Auge besitzt zwei Achsen, die unter einem Winkel von 5 – 7° zueinander geneigt sind: eine sog. Augenachse und eine Sehachse. Die biaxiale Anlage des Auges wird ergänzt durch eine bipolare Konstruktion des Zentralgebiets. Die differenzierte Besetzung der Retina mit spezifischen Sensoren und deren abgestimmtes Zusammenwirken ist am Einfachsten vorstellbar – sowohl evolutionär, ontogenetisch als auch hinsichtlich eines aktuellen Wahrnehmungsaktes – als Summe und Ergebnis selbstregulierender Vorgänge
in Abhängigkeit von den Lichtverhältnissen und Sehleistungen. Die Verteilung der Zapfenstandorte entspricht einem geometrischen Evolventen-Muster aus zwei gegeneinander verschränkten Spiralen mit peripherer Zunahme der sie umgebenden Stäbchenanzahl. In deren gitteroptischer Interpretation erweist sich nach Lauinger, dass in den Körnerschichten der Retina ganz bestimmte Funktionalitäten bereits optisch erarbeitet werden, dass zelluläre optische Gitternetze und optische Säulen die Information bereits pixelübergreifend vorstrukturieren, also ein einzelnes Ganzes separieren können. Danach würde eine schichtenweise visuelle Informationsbearbeitung bereits in der invertierten Netzhaut einsetzen.
In Abb. 20 sind lokale gitteroptische Metapixel dargestellt. Konzentrische RGB-Beugungs- ordnungen im Fresnel-Nahfeld hinter den zellulären diffraktiven Gittern stehen lokalen Gruppen von Fotorezeptoren zur Verfügung. Die rechte Abb. zeigt eine Aufsicht auf die interferenzoptische Struktur der Fresnel-Ebene hinter zwei hexagonalen Gittern (Gitterkonstante im 1. Gitter ca. 80μm, im 2. Gitter ca. 20μm) [Lauinger 2014]
Lokale gitteroptische Metapixel rechts: Interferenzoptische Struktur der Fresnel-Ebene hinter zwei Gittern
unten links: Fraunhofer-Beugungs- bild im Fernfeld hinter einem hexagonalen Gitter bei Verwendung von weißem Licht.
unten rechts: Die weiße 0. Beugungsordnung liegt im Zentrum und die chromatischen 1. u. 2. Ordnung liegen in Schnittpunkten zwischen versetzten Parallelen- u. Hyperbel- scharen sowie auf Kreisen um den Mittelpunkt. (Lauinger 2014)
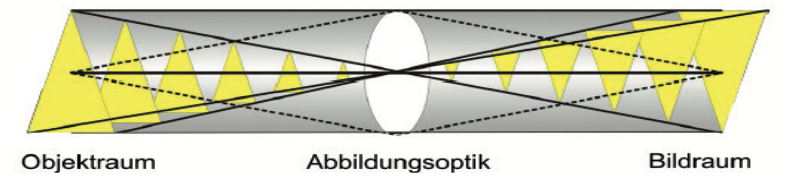
Fragestellung 7: Inwieweit wird unser Sehen durch holistische und synchronisierende Ansprüche nach Ganzheit bestimmt?
Lauinger hat zu dieser Frage einige Aspekte neu ins Feld geführt: „Die geometrische Abbildungsoptik überträgt jedes Objekt als Ganzes in einer globalen optischen Säule, einem achsenzentrierten Rotationskörper, in sein Bild im Bildraum des Auges. Ein sichtbares Objekt wird also ‚holistisch‘ – als Ganzes – übertragen. Das einzelne Objekt ist somit zuallererst optisch ein Ganzes.
Stehende optische Interferenzfelder ordnen eine Vielzahl von zellulären Elementen zu quasi- kristallinen hexagonalen Dichtestpackungen und bewirken ein ‚optical binding‘ der Zellen im elektromagnetischen Feld, wie Burns et al. …eindrücklich demonstriert haben. Es gibt genügend Hinweise, dass auch – trotz schichten – und säulenweiser neuronaler Weiterverarbeitung – in der Netzhaut und in visuellen Zentren des Gehirns ein Objekt weiterhin als Ganzes zusammengehalten wird. Dem ‚optical binding‘ kann so unmittelbar das ‚neuronal binding‘ folgen. Prominente Gehirnforscher stellen ein „Feuern im Gleichtakt“ der Nervenzellen dann – und nur dann – fest, „wenn das dargebotene Muster optisch wie ein einziges Objekt erscheint“. Neuronen im visuellen Cortex von Katzen synchronisieren ihre Oszillationen über räumlich getrennte Bereiche, „wenn Neuronengruppen durch Reize stimuliert werden, die gemeinsame Eigenschaften haben, z.B. alle zu einer zusammenhängenden Figur gehören“ [Singer 1990] Durch die Synchronisation in gekoppelten harmonischen Oszillationen bleibt ein Objekt mit seinen Teilen auch in den kortikalen Säulen und weit auseinanderliegenden Rezeptiven Feldern des Gehirns als synchrones Ganzes erhalten. [Lauinger 2014, S. 262]
Auch Krumeich u. Knülle-Wenzel gelangten zur Annahme, dass ihre musterartigen ‚Colorgramme‘ als Ganzes synchron übertragen werden können: „Das Bild der Umgebung, das spiegelartig zur Verfügung steht, wird…reflektiert….Wir glauben, dass dieser Prozess mit einem Spiegel vergleichbar ist, der vor uns bewegt wird, oder ähnlich, dass wir vor dem Spiegel bewegt werden und somit ein ..reflektiertes Bild wahrnehmen.“ Diese Vorstellung korrespondiert mit der Annahme von Funk, dass unsere Augen wie Scanner funktionieren könnten, die das Blickfeld abtasten.
Auch die auf das Symmetrieprinzip sich stützende ‚Farbgestalt-Hypothese‘ des Autors geht in ähnlicher Weise davon aus, dass sich das aufgenommene elektromagnetische Reizprofil unter Mitwirkung aller Schwingungsebenen jeweils als Ganzes gestalthaft in eine sinnesphysiologische Synchronisation einbringt Abb. [Bendin 1991].
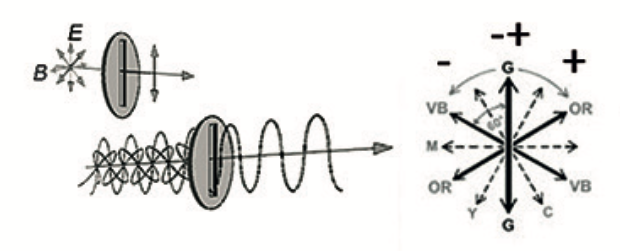
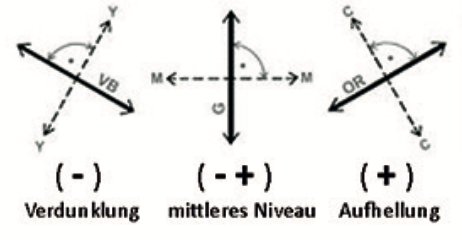
Abb. 22: Symmetrisches Wirkprinzip nach Bendin 1991
Fragestellung 8: Erlauben die neuen Ansätze auch eine erweiterte Interpretation der bisher angenommenen örtlichen Aufteilung der Signalverarbeitung nach verschiedenen Qualitäten des Sehens (Farb-, Form-, Tiefen- u. Bewegungssehen)?
Wenn in der invertierten Retina des Auges eine optische Vorstrukturierung des Sichtbaren und eine interferenzoptische Informations-Vorverarbeitung stattfindet und auf ihr aufbau- end eine schichtenweise neuronale Weiterverarbeitung, dann müssen beide aufeinander abgestimmt und miteinander verschränkt sein. Da derzeit aber die Art der Verknüpfung der optischen mit einer – bisher nur in Teilen aufgeklärten – neuronalen Vorstrukturierung u. -verarbeitung nur in Ansätzen beschreibbar ist, wird es weiterer Forschungsarbeit bedürfen, das Zusammenwirken gezielt zu untersuchen. Nach der Einschätzung von Funk z.B. zeigt sich, „dass die bisher üblichen Methoden noch lange nicht subtil genug sind, um eventuelle beteiligte elektrische, quantenmechanische oder holographische …Phänomene in den neuronalen Netzwerken zu erfassen.“ [Funk 2016] Wie sich aber in einigen neuen Denkansätzen 100 Jahre nach Schopenhauers Schrift ‚Über das Sehn und die Farben‘ bereits gezeigt hat, sind alternative Erklärungsmodelle zu den bisherigen Annahmen nicht nur vorstellbar, sondern könnten durchaus zum Überdenken auch der bislang allgemein noch angenommenen Separierung des Farb-, Form-, Tiefen- u. Bewegungs-Sehens führen. Insbesondere der Kern der SWA-Theorie [Krumeich/Knülle 1992], dass durch Reflexion des Lichtes an der Bruchschen Membran die Außenglieder der Rezeptoren stehende Wellen direkt aufnehmen und in Muster (Colorgramme) umwandeln könnten, hätte im Zusammenwirken mit der Aufdeckung der Funktion der Müllerzellen als Lichtleiter [Franze et.al. 2007] und dem einleuchtenden gitteroptischen Drei-Schichtenmodells auf der Grundlage der biaxial und bipolar ausgerichteten Rezeptorverteilung [Lauinger 1994/96/2014] das Potential zu neuen Einsichten in die inverse Funktion der Retina. Möglicherweise könnte dies auch zu einem Korrektiv werden für bisherige Interpretationen der nachgeschalteten Signalverarbeitungen in der Netzhaut, im CGL und in der Area V1. Angesichts der evolutionär begründeten, fortwährend aber auch ontogenetischen Selbstorganisation während der plastischen Lernphase des Sehens erhielte dann auch Wilhelm Wundts frühe Vision zu Art und Struktur der Rezeptoren als „einer aus unbestimmt vielen Abstufungen bestehen- de(n) Reihe. [Wundt 1910] “ einen neuen Stellenwert. All dies legt strukturelle Analogien zwischen den Reizkonfigurationen als ‚Erkanntem‘ und den Verarbeitungsstrukturen als dem ‚Erkennenden‘ nahe entsprechend der Prämisse der Erkenntnislehre des Empedokles ( 492-432 v. Chr. ): „Gleiches wird durch Gleiches erkannt.“ Dass jene Entsprechungen aus biologischen Sicht stets auch selektiver Dynamik unterliegen, machte Andrè Karlicek in seinem Einführungsbeitrag deutlich: „Ein ernsthafter Diskurs über Farben kann auch in Zukunft nur auf der Basis einer kritischen Ästhetik geführt werden, die dann aber gleichermaßen ontogenetische und evolutive Elemente zu berücksichtigen hat.“[Karliczek 2016]
Vollständiger Beitrag zum Download:
Verantwortung liegt bei dem Urheber des Beitrags: Eckard Bendin

